There are many parallels between how the collective intelligence of neurons and the collective intelligence of other cells navigates behavioral and morphogenetic spaces respectively. This is discussed in detail for example here and here. Biological systems at many scales navigate these spaces. But they seem like such different worlds – the familiar 3-dimensional arena of movement, vs. the high-dimensional space of possible anatomical configurations.
Here I briefly mention a few places where these systems intersect. One interesting example is Physarum – a slime mold that solves problems by growing in different patterns. Here it is using biomechanics to detect masses at a distance and reliably choose to grow towards the bigger strain angle:
This was work done by Nirosha Murugan and others in my lab. It’s an inexpensive model system for a lot of fascinating questions in basal cognition. My favorite part is that until ~800 minutes, it hasn’t actually made a decision – it’s gathering data, by rhythmically tugging gently on the gelatinous substrate to get information about the distribution of mass in the vicinity:
I have no idea what the ecological significance of this might be in the wild.
In this case, the Physarum’s behavioral space is its morphogenetic space: control of its body shape is how it navigates its environment. Another example of crossing spaces can be seen for example in bacteria: a given cell might move up a sugar concentration gradient, to receive more nutrients, or, transcribe a new enzyme to be able to exploit a different sugar instead. In that case, the options for the cell combine transcriptional and conventional 3D spaces. I have argued before that these distinctions into different problem spaces are specific perspectives by external observers (e.g., us) and may not at all be how the agent itself perceives its option landscape.
Now, let’s look at some interesting examples of how animals connect behavioral and morphological controls. This is surprising to many, but actually you do not need a brain to develop – most morphogenetic processes happen fine without a brain (and indeed many happen before there is any brain). However, there are interesting examples where the presence (and possibly activity) of the nervous system makes important impacts on morphogenesis.
First, it’s been known for a long time that regeneration requires innervation. But, oddly, this is not fundamental but acquired: amphibians which developed without the relevant nerves were also able to regenerate their limbs without it, giving rise to the idea that some body structures literally become addicted to nerves.
Then, there are interesting examples such as this:

What is shown here are the results of cutting and deviating a central nerve (the color additions to this figure, from Regeneration in Animals and Related Problems, edited by V. K. a. H. A. L. Trampusch, are mine). When deviated to a wound on the side of the organism, the backward-facing end of the nerve induces a tail, while the anterior-facing end of the nerve induces a head. This connection between direction and anatomical identity is also found in planaria, in this and this work led by Johanna Bischof. It’s a fascinating and still largely-untapped area of research that connects subcellular directional polarity with organ identity and whole organism-scale anatomical polarity, as we also found in the patterning of the left-right axis during development.
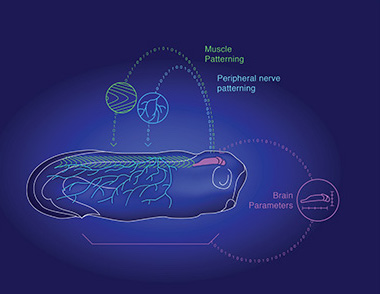
In frog embryos, we found that most of the embryo develops fine without a brain, except that both muscle and peripheral nerve are disrupted. Regions in the middle and posterior of the animal require something about the presence of the brain in order to form properly. Remarkably, it turns out that brainless embryos are more sensitive to teratogens: in other words, the presence of the brain helps protect the embryo from various disruptors of morphogenesis. The brain joins CEMA (cross-embryo morphogenetic assistance) as one of the many positive influences that complicate the question of “how harmful is a given drug?”, because what we see is not raw teratogenicity, but teratogenicity after it has been ameliorated by a number of influences from within and without.
Interestingly, the effects of a missing brain can be largely erased by the misexpression of a specific ion channel – HCN2 – which we previously showed can also repair a variety of other birth defects.

This is described in detail here and here. Also, the innate immunity of the tadpole is abnormal without the presence of the brain, and immune-bacteria interaction is significantly impacted by the presence or absence of brain.
While there are many distributed intelligences in the body that are not neural, the brain has many still-undiscovered roles. And, those roles begin very early in development – long before movement and “behavior” appears.
Leave a Reply to Robert Stern Cancel reply