The typical way to think about embryogenesis (also metamorphosis, regeneration, etc.) is that a group of cells cooperate toward creating and maintaining a specific anatomical outcome (also known as a Target Morphology for a given species). This requires navigating the space of all possible geometries, starting from an initial state (e.g., fertilized egg cell) and ending up in a specific region of that anatomical morphospace. Importantly, this is not simply an issue of emergent complexity, inexorably rolling down a prescribed path (because morphogenesis has remarkable competencies to adjust for novel circumstances to reach their goal state – just like autonomous vehicles, some animals, etc.).
One interesting question that can be asked about this process is: where does all the information come from – how does the collective know where in morphospace to go (what shape to build)? The typical answer is that it comes from the genome and the chemical products placed into the egg by the maternal organism – that is, it is passed down vertically and each embryo has everything it needs to complete its journey. Frogs, fish, and many other non-mammals do this outside of the maternal organism, and we tend to think of the egg as a self-contained packet of information and resources needed for morphogenesis. Angela Tung (a PhD student in my lab) and I began by reviewing interesting exceptions to this idea – cases where important information and influence arrives “laterally”, not passed down via the zygotic genome and maternal egg products.
Another question you might ask is: what about scale-free/multiscale dynamics – symmetries in how things happen on small and larger scales (e.g., “as above so below”): if chemical networks work together to build cells, and cells cooperate to build the correct body, is that where it stops for developmental biology, or could there be a higher level? My multi-scale perspective predicts that there should be higher levels. For example, that multiple embryos could form a sort of “hyper-embryo”, with its own new competencies and its own path through gene expression and physiological spaces. We tested this unconventional hypothesis and found some very interesting results. This post discusses our paper that just came out on this topic, originally preprinted here and spearheaded by Angela Tung and involving a team of graduate students, undergraduates, and post-docs. The official paper is here, and we discuss it on Curt Jaimungal’s podcast in this episode.
Some fundamental observations in this paper are:
- Large groups of frog embryos are much more resistant to teratogens (agents that disrupt morphogenesis by confusing cell signaling) than are small groups or embryos cultured alone. This is true even though we scale the teratogen appropriately to large groups (making sure that no matter the group size, each embryo gets the same amount of drug), and it’s also true when we use molecular-genetic stressors (mutant mRNA injections instead of drugs) to target very different pathways – it’s not about drug metabolism, dilutions, or any one specific signaling pathway. We call this effect CEMA – Cross Embryo Morphogenetic Assistance.
- It’s not just the Wisdom of Crowds phenomenon. Our initial simple model was that every teratogen-exposed embryo is confused about a random part of the information it needs, and in a crowd, they all fill in each other’s informational gaps. That would predict that a large group made up of half stressed embryos and half embryos that never saw the teratogen should do very well, because the pristine embryos should have all the correct information and would be able to instruct their siblings better than ones that had also been stressed could do. In fact what we saw is that it only works if every member of the large group has seen the teratogen. Something more complex is going on, where you have to have had the problem, in order to help others overcome it. We included an agent-based model of embryonic noise and signaling to try to explain the dynamics.
- We narrowed down the mechanism by 3D-printing tiny little “jail cells” for each embryo so that large groups could be in the same dish of medium but couldn’t touch each other, and similar methods that tested various ways that embryos could use to signal among the group. The bottom line is that it looks like the signal is short-range chemical communication, apparently involving the calcium ion and ATP.
- We did unbiased transcriptomic profiling, asking which genes are turned on by large vs. small groups responding to the same challenge. We found numerous genes that only activate in large groups, which gives an entrypoint into the downstream mechanisms of CEMA activated after those ‘group repair’ genes are turned on. We found some genes that are specific to large groups (unstressed, normal development) and other genes that are only turned on in large groups (but not small ones) challenged by teratogen exposure. Thus, the hyper-embryo has its own transcriptome that we can now begin to unravel.
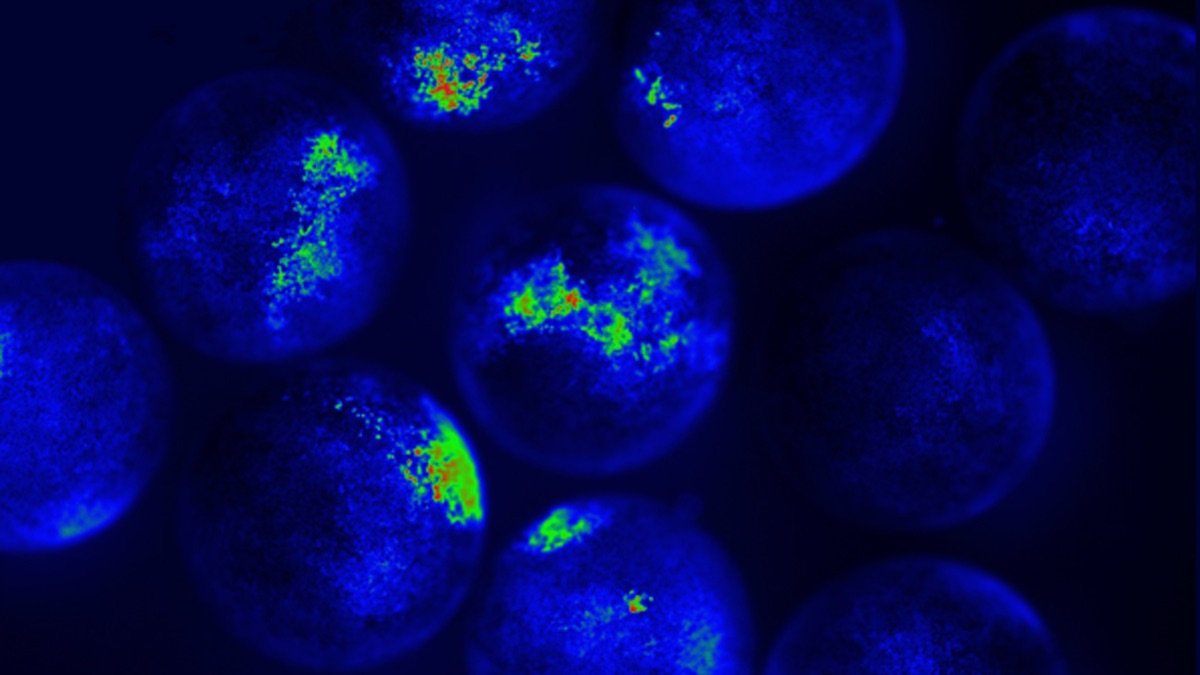
- We developed an assay to watch the embryos communicate. It’s quite amazing – by tracking calcium signaling (a sort of generic read-out of cellular computations) in embryos that were poked, we can see that the information propagates not only across an embryo, but also between embryos. This is something that has been seen within cells and tissues, by one of the key figures in modern bioelectricity (Lionel Jaffe, his early paper in this book) and modern workers such as here, here, and here. Calcium is a nice generic read-out of cellular information processing. Here is a typical video, where you can see the injury wave within the poked (left) embryo and after a but of time, the right embryo responds:
Here’s another amazing video that Patrick McMillen made a while back, showing information propagating across individual Xenopus tissues separated by aqueous medium, after the left one gets poked:
Some major questions are being addressed next:
- What are the rest of the mechanistic steps in the communication – besides calcium and ATP, what else is involved?
- How does the encoding work? Complex information on how to build an embryo head cannot be encoded in something as flat as the local concentration of a signaling molecule in the water. There needs to be more bandwidth; how does it work – is it patterned (in time and/or space) in the medium? These molecules are hard to track directly, they are too small to have tracers put on them without drastically changing their properties. But we have some ideas about how to test the information carrying capacity of this kind of effect.
- How widely does CEMA extend – what is the range of genetic and biophysical perturbations that it can assist with, and does it work in mammals (i.e., biomedical assays)?
- Will it work across species, or is it only a within-species effect?
We will soon find out. The big opportunity here is to begin to develop a kind of “hyper-developmental biology”, which seeks to work out the rules by which embryos work together to achieve robustness at the population level (paralleling the standard developmental biology research program which operates at a lower scale). But there are a number of key implications:
- Tens of thousands of papers that report the teratogenicity level of some chemical or a mutation are in fact reporting the corrected value – the effect size after any CEMA-like processes have exerted corrective influence. This means we don’t really know the true effect size of many of these things, and that future papers have to pay much closer attention to the cohort size when reporting such data. Besides the impact on the published literature, there is a deep issue here which relates to within-embryo repair (regulative competency) as well: evolutionary selection, like scientists, sees adjusted phenotypic fitness after corrections have been made. For us, this results in an under-appreciation of the potency of our perturbations. For evolution, it results in an intelligence ratchet in which subsequent advances target the repair machinery more than the structural genome (see this paper).
- What if we could artificially fake CEMA? What if, for a singleton human patient, we could provide whatever signals one might normally get from a group of organisms, to help healing, or resistance of aging/cancer/degenerative disease?
- More fundamentally, what does this mean for the genotype-phenotype map? In other words: the amazing robustness of development is not simply due to the vertical competencies of the genomically-provided hardware. Important aspects seem to be lateral, a function of the parallelization of the physiological software running on many embryos at the same time and sharing information as a collective.
And finally, here are some broader thoughts on this kind of result. It fits in very well with our development of the TAME framework of multiscale agency; as I have argued, morphogenesis is a cellular collective intelligence solving problems in morphospace, and it appears that groups of embryos meet the same criterion. Note that our RNAseq (transcriptional profiling) data revealed specific genes that are only expressed when CEMA-capable large group, not small groups, are confronted with teratogens. This may be the first study of the unique mRNA expressed in a hyper-embryo with and without developmental insult. Moreover, given my proposal that morphogenesis and regeneration are basically the behavior of a collective intelligence of cells in anatomical morphospace, we can also consider that the drug we used to perturb embryo:embryo signaling (and abolish the CEMA effect) is basically a general anesthetic for the morphogenetic group mind of the large cohort. This parallels the way in which conventional anesthetics prevent bioelectrical communication between cells (especially neurons), leading to the temporary disappearance of the high-level agent and of its problem-solving capabilities (in our case, top-down morphogenetic problem-solving competencies). This is a very testable prediction, using the work of G. Tononi, Erik Hoel, and others working on new information theory for detection of emergence of new higher-level entities: we will soon be processing physiological data of from groups vs. singleton embryos to quantitatively detect emergence of a kind of virtual governor whose body is the entire group.
There are many papers that can now be seen to be relevant to the CEMA topic in ecology, plant biology, microbial biofilms, etc. Even stem cell biology has relevant phenomena, where many cell types simply do not survive alone in a dish, even though their metabolic needs are provided for. It’s nice to see that vertebrate embryogenesis does not escape the symmetry of collective intelligence. The relevance to multi-human groups (e.g., social buffering of medical states or Kangaroo Care of human infants) remains to be explored. My guess is that there will be deep connections to neuroscience and to emerging theories that tie together evolutionary and cognitive change.
Leave a Reply to Benjamin L Cancel reply